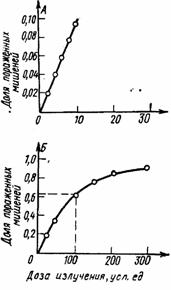
Рис. III.35. Зависимость изменения эффекта от дозы облучения. А и Б — при малых и больших дозах соответственно. Рассмотрим определение основного параметра радиочувствительности - величину d0 или d37, которую в теории мишени принято называть инактивирующей, или среднелетальной (не путать с ЛД50) дозой, на примере анализа одноударного поражения. В качестве такового подразумевают гибель облучаемого объекта от эффективного единичного попадания в мишень.Тогда если N0 — исходное число объектов, а N — число объектов, не пораженных излучением при дозе d, то выход из строя определенной доли объектов dn/n при приращении дозы dd выражается уравнением dN/N = -dD/d0, где d0 - доза, при которой на каждый объект в среднем приходится одно попадание (отсюда и наименование d0 - среднелетальная доза).
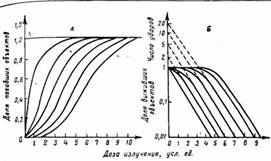
Рис. III.36. Дозовые кривые выживания для объектов с различной ударностью мишени. А - в обычном масштабе (S-образные, или сигмоидные, кривые; число ударов обозначено на кривых); Б—в полулогарифмическом масштабе
При интегрировании данного уравнения получаем
![]() ,
,
где е — основание натуральных логарифмов.В природе чаще встречается другой тип кривых, характерный для большинства клеток растительного и животного происхождения; примеры таких кривых были уже приведены ранее. В линейных координатах они имеют S-образную форму (рис. III.36, А). В этих случаях говорят о многоударном процессе, имея в виду, что для инактивации объекта необходимо не одно, а два и более попаданий в единственную мишень или поражение двух мишеней и более, каждая из которых должна быть поражена.При изображении таких кривых в полулогарифмическом масштабе (рис. III.36, Б) они приобретают плечо, переходящее в прямолинейный участок, наклон которого совпадает с наклоном соответствующей одноударной кривой. Экстраполированием прямолинейного участка кривой к нулевой дозе на оси ординат отсекаются отрезки, соответствующие «ударности» мишеней (или их числу). Изображение многоударных кривых в полулогарифмическом масштабе позволяет достаточно точно оценить выживаемость при ее малых значениях (больших дозах) и легко определить экстраполяционное число, характеризующее число мишеней (ударов), поражение которых необходимо для инактивации клеток.Нужно иметь в виду, что возможность строгого применения теории мишени уже в самом начале была ограничена ее основоположниками (Д. Ли, 1946) областью анализа одноударных эффектов. Интерпретация конкретных многоударных кривых с тех же позиций затруднительна. Прежде всего, это связано с многочисленными экспериментальными фактами изменения экстраполяционного числа при применении самых различных модифицирующих агентов или изменении условий жизнедеятельности объектов, что само по себе не должно сказываться на числе мишеней. Кроме того, экстраполяционное число в ряде случаев достигает десятков и сотен единиц, что не позволяет их отождествить с числом мишеней в клетке.По мере развития экспериментального изучения репарации потенциальных повреждений размер плеча на кривой выживания стали связывать со способностью клеток к пострадиационному восстановлению, а величиной плеча характеризовать их репарационную способность. Однако и в этом случае возникают большие трудности при попытках интерпретации, с позиций классической теории мишени, многих клеточных радиационных эффектов, обусловленных, например, изменением химизма клетки в результате радиолиза ее жидких компонентов или метаболического взаимодействия различных клеточных органелл. Эта теория не касается природы мишеней, ответственных за гибель клетки. Такими мишенями могут быть уникальные макромолекулы, определенные участки мембран и другие структуры, попадание в которые приводит к поражению. Поэтому для определения природы мишени следует привлекать данные, получаемые при использовании специальных методов исследования. Именно эти обстоятельства уже в 40-ых годах сделали очевидным необходимость строгого ограничения применения теории мишени только для анализа поражения одноударных эффектов.В частности, потребовалось расширить само понятие попадания, так как стало известно, что первичные повреждения элементарных клеточных структур могут вызываться не только непосредственной их ионизацией, но и опосредованной: химически активными продуктами радиолиза окружающей среды. Кроме того, вследствие обнаружения миграции энергии по макромолекулам даже при непосредственных формах взаимодействия, реализация повреждения может происходить далеко от места первоначального «попадания».Таким образом, классическая теория попаданий, плодотворная при анализе количественных закономерностей радиационного поражения отдельных мишеней, не ставит своей задачей решения вопроса о том, какими процессами связано поражение мишеней с проявлением конечного эффекта.В основе теории мишени лежало предположение о том, что характерные зависимости доза—эффект могут быть отражением квантованного характера взаимодействия излучения с веществом и наличия в клетках высокочувствительных объемов — мишеней.В случае очень массивных повреждений конечный эффект действительно детерминирован уже на начальной стадии, но в подавляющем большинстве других ситуаций все случайные факторы затрагивают более поздние звенья причинно-следственной цепи, ведущей к конечному эффекту. При этом каждому лабильному состоянию объекта соответствует определенная повышенная вероятность наступления регистрируемой реакции, возрастающая с увеличением предварительного повреждения. Таким образом, при формировании радиобиологического эффекта всегда существует суперпозиция множества случайных событий, что и учитывает стохастическое рассмотрение.При таком подходе любой биологический объект, в частности клетку, представляют в качестве лабильной динамической системы, которая постоянно находится в стадии перехода из одного состояния в другое. Вследствие крайней сложности системы любой такой переход сопровождается и связан с множеством комплексных и элементарных сопряженных реакций отдельных клеточных органелл и макромолекул. Естественно, что в процессе жизнедеятельности, благодаря влиянию самых разнообразных, не подлежащих учету факторов и малейших сдвигов в исходном состоянии, возникает вероятность «отказов» в элементарных звеньях, а вследствие этого — «крушения» всей системы. Поэтому любое критическое событие, например митоз или гибель клетки, можно предсказать лишь с известной вероятностью.На биологическую стохастичность при облучении объекта накладывается стохастичность вследствие случайного характера взаимодействия излучения с веществом, что резко увеличивает вероятность «крушений» системы, происходящих со значительно меньшей частотой и в необлученном контроле.Стохастическая теория рассматривает различные возмущения биологической системы, возникающие в процессе жизнедеятельности или под влиянием облучения, с позиций теории вероятностей, стремясь описать их моделями, максимально соответствующими представлениям динамической биохимии и молекулярной радиобиологии. В этом случае мишенями являются все компоненты живой системы, а регистрируемая реакция обусловлена суперпозицией самых разных событий.Существенно, что стохастическая гипотеза учитывает как физиологические, так и индуцированные излучением процессы в их динамике, в то время как классическая теория мишени рассматривает эффекты, вызванные облучением, как строго детерминированные первичными актами абсорбции энергии.Используя аппарат стохастической гипотезы, можно учесть реальное взаимодействие ряда последовательных попаданий, а также влияние фактора времени, репарационных процессов, роль ЛПЭ и т.д. Все это может быть выражено системой дифференциальных уравнений, описывающих переход биологического объекта под влиянием облучения из одного состояния в другое. В принципе можно учесть количественно влияние любого модифицирующего фактора на соответствующие дозовые зависимости.Более того, этот аппарат может быть использован для анализа не только кинетических процессов на молекулярном уровне, но и морфологических процессов, образования новой клетки, ее дифференцировки и др.Таким образом, « . дискретное изображение непрерывных физиологичесих процессов не является приближением, которое принимают только ради удобств, а становится необходимым при изучении различного поведения отдельных единиц популяции . Действие излучения на клетку свободно укладывается в эту общую схему, что также выражается в дискретных событиях. При этом не существенно, происходят ли соответствующие критические события в результате дискретных актов абсорбции энергии или в результате усиленной облучением лабильности, органически свойственной биологической системе . В этой схеме можно учесть даже такие сложные явления, как феномены обратной связи и механизмы регуляции, т. е. жизненные процессы, которые выявляются в результате нарушений, если экспериментальные данные указывают на необходимость и возможность такого учета» Эффект, обусловленный попаданием в одну из уникальных структур клетки, приводящим ее к гибели, формально может быть описан как с классических позиций теории мишени, так и с позиций стохастической теории. Иными словами, выводы теории мишени являются частным случаем стохастического подхода.Проведенные обсчеты некоторых экспериментальных результатов с помощь аппарата стохастической теории показали, что для инактивации клеток млекопитающих редко ионизирующим излучением в среднем необходимо совместное действие не менее четырех событий абсорбции.Итак, в соответствии с основными исходными позициями стохастическая концепция предлагает как бы более «биологическую» интерпретацию кривых доза — эффект по сравнению с их объяснением с позиций теории мишени, хотя основное положение последней о том, что эти кривые определяются в основном случайной природой абсорбции энергии, остается незыблемым.Для обозначения клеточных повреждений, которые нельзя отождествлять с локальными изменениями клеточных структур, прежде всего генетических, стохастическая теория вводит понятие «дисперсного начального повреждения». Природа такого повреждения весьма разнообразна: изменение клеточных мембран и других множественных структур, инактивация какой-либо жизненно важной системы и др.Репаративные процессы, сказывающиеся на конечном эффекте, учитываются понятием компенсационной способности объекта. При анализе дозовых кривых с учетом функциональной лабильности биологических объектов стохастическая гипотеза приводит к пониманию того, что экспоненциальная кривая указывает на систему без компенсаторных механизмов, а сигмоидная — соответствует системам, обладающим такими механизмами, эффективность которых снижается при возрастании дозы облучения. Экстраполяционное число же следует рассматривать как количественное выражение компенсационной способности облучаемого объекта, а не как «число мишеней».Таким образом, сама по себе экспоненциальная зависимость эффекта от дозы далеко не предопределяет решение о наличии первичных одно- или многоударных реакций, а с полным основанием делает логичной другую интерпретацию.Подводя итоги изложения обеих концепций — классической теории мишени и стохастической теории, необходимо, прежде всего, подчеркнуть их основное общее свойство — строго количественный подход. Более того, стохастическая теория представляет собой логическое дополнение классических представлений, которые не могли объяснить многообразие радиобиологических феноменов, да и не претендовали на это.Вполне естественно, что первоначальные гипотезы исходили из упрощенных представлений о механизме первичных радиобиологических процессов на основе чисто физических, а позднее радиационно-химических закономерностей, установленных при облучении простых систем. Однако именно эти простые, общие и формальные схемы впервые перевели радиобиологию с описательных позиций на прочную количественную основу. Круг возможного применения первоначальных представлений был очерчен еще при их формировании, и в этом плане были получены наиболее значительные результаты. Дальнейшее накопление новых фактов потребовало развития более широких подходов.Нельзя не заметить, что при этом остались незыблемыми оба определяющих фактора классической теории мишеней — дискретность радиационного агента и функциональная негомогенность биологического объекта. Существенно, однако, что если в теории мишеней последняя определяется наличием фиксированных мишеней, поражение которых уже определяет конечный эффект, то в системе новых представлений показана несостоятельность такой точки зрения, взамен которой развита идея определяющей роли стохастической природы физиологических процессов и их радиационных нарушений.Стохастическая гипотеза учитывает современные данные о микрораспределении энергии, вариабельности радиочувствительности логических объектов, а также роль репарационных процессов. Тем самым она значительно расширяет круг явлений, которые можно интерпретировать с новых позиций. Она более «биологична» по своей природе, так как устанавливает связи с конкретными морфологическими и функциональными разделами цитологии и физиологии. Однако ее математический аппарат достаточно сложен и это затрудняет ее широкое применение.Анализ количественных закономерностей различных реакций дрожжевых клеток на облучение показал, что ни классические модели, основанные на принципе попадания, ни концепция биологической стохастичности не способны объяснить всю совокупность полученных экспериментальных данных. Дополнив принцип попадания предположением о вероятностном характере проявления повреждений,)предложил вероятностную модель радиационного поражения клетки.Согласно этой модели разные клетки, подвергнутые облучению в одной и той же дозе, поражаются в разной степени в соответствии с принципом попадания, но в отличие от классических представлений и потенциальные, и реализованные повреждения проявляются с вероятностью меньше единицы. Реализованные повреждения (или индуцированные ими изменения) наследуются при делении клеток и с некоторой вероятностью, зависящей от числа этих повреждений, приводят к неосуществлению клеточного деления. При этом вероятность проявления повреждения может зависеть как от биологических (генетических) особенностей клеток, так и от условий их культивирования, увеличиваясь при ухудшении этих условий.Вегетативное размножение облученных клеток согласно этой модели - случайный процесс «рождения и гибели», приводящий к появлению нежизнеспособных клеток в отдельных размножающихся клонах (т. е. к наблюдающейся в эксперименте частичной инактивации); дело случая, образует ли данная облученная клетка макроколонию или погибнет после того или иного числа делений.Таким образом, формально вероятностная модель, являясь как бы синтезом принципа попадания и стохастической концепции, используя сохранившие значение основные положения первого, дополняет последнюю, расширяя диапазон явлений, которые могут быть интерпретированы с позиций биологической стохастичности. Описывая не только качественно, но и количественно (с помощью специальной системы уравнений) различные проявления нарушений репродуктивной способности клеток, вероятностная модель выгодно отличается от классических схем тем, что позволяет предсказать наблюдаемое многообразие радиобиологических эффектов на основе минимума исходных предположений.Тем не менее, вероятностная модель, в значительной степени основанная на принципе попадания, имеет много общего с классическими моделями. В тех случаях, когда вероятность проявления повреждения равна единице, вероятностная модель переходит в классическую одноударную модель. При качественном анализе экспериментальных данных, который ограничивается средними характеристиками популяции облученных клеток, вероятностная модель зачастую приводит к таким же результатам, как и многоударная.Главное отличие вероятностной модели от классических состоит в том, что согласно последним радиочувствительность клетки определяется лишь объемом мишени и критическим числом попаданий. С позиций же вероятностной модели проблема радиочувствительности представляется более сложной. Процесс радиационного поражения клетки Ю. Г. Капульцевич формально делит на три этапа.Первый этап радиационного поражения - осуществление событий попадания, в результате которых формируются первичные потенциальные повреждения. Вероятность образования первичного повреждения на единицу дозы облучения зависит от величины эффективного объема и от величины энергии, необходимой для образования первичного повреждения. Поскольку отнюдь не всякое выделение энергии в эффективном объеме может привести к образованию первичного повреждения, клетка, по-видимому, способна восстанавливаться еще на стадии формирования потенциальных повреждений. На этой стадии радиочувствительность определяется величиной эффективного объема (V) и вероятностью (р) образования потенциального повреждения (при локальном выделении энергии в эффективном объеме). Параметры V и р могут зависеть как от биологических особенностей объекта, так и от условий облучения, например от температуры, влажности, концентрации кислорода и др.Второй этап радиационного поражения — реализация потенциальных повреждений. Так как клетки способны восстанавливаться от лучевых повреждений, то реализованными оказываются не все возникшие потенциальные повреждения, а лишь часть их (r). Следовательно, радиочувствительность клетки определяется и вероятностью реализации потенциального повреждения. Все три параметра (V, р и r) определяют частоту (b) реализованных повреждений на единицу дозы облучения (b = Vpr) и среднее число α реализованных повреждений при дозе D, т. е. α = bD. С помощью параметра b вероятностная модель учитывает зависимость радиобиологических эффектов от дозы облучения, ЛПЭ и продолжительности восстановления.Третий этап радиационного поражения — различные вторичные нарушения нормального протекания внутриклеточных процессов, вызываемые реализацией повреждений. По-видимому, и на этом этапе возможно восстановление клеток от последствий реализованных повреждений или их компенсация, поэтому вероятность проявления реализованного повреждения не равна единице, а характеризуется величиной α, которая зависит от биологических особенностей клетки и от условий культивирования. Чем больше надежность системы, определяемая способностью клетки устранять всякие отклонения от нормы в работе внутриклеточных структур, тем меньше величина α. С помощью параметра α вероятностная модель позволяет анализировать пострадиационные модификации эффектов, не связанные с восстановлением.Таким образом, радиочувствительность клетки, о которой судят по кривой выживания, определяется четырьмя параметрами: V, р, r, α. Эти параметры достаточны не только для описания формы кривой, но и всего многообразия проявления действия излучений на репродуктивную способность клеток, зависимость радиобиологических эффектов от условий культивирования клеток и физических характеристик излучения.Таковы основные качества вероятностной модели, которую можно считать логическим звеном в развитии количественной радиобиологии.Однако ни сама модель, ни производимый с ее помощью анализ реакций клеток на облучение не позволяют выявить природу повреждений, лежащих в основе этих реакций. Кроме того, нельзя забывать, что сделанные выводы справедливы только для дрожжевых клеток, что затрудняет проверку применимости вероятностной модели к описанию лучевой реакции клеток млекопитающих.Ниже будут рассмотрены качественные концепции, характерная черта которых заключается в попытке указать конкретные биохимические процессы, определяющие биологическое действие радиации. Основная роль при этом отводится высокореакционноспособным продуктам — радиотоксинам,образующимся в биосубстрате вслед за поглощением энергии излучения и инициирующим множественные повреждения различных клеточных органелл.Радиобиологический парадокс несоответствия малых количеств поглощенной энергии с разительным биологическим эффектом с позиций качественных гипотез объясняется наличием различных биологических механизмов усиления первичных поражений.Еще А. С. Мочалиной и Ю. Б. Кудряшовым в лаборатории Б. Н. Тарусова было обнаружено, что водно-солевые вытяжки из облученной печени при введении интактным животным вызывают гемолиз. В связи с этим было высказано предположение, что под влиянием облучения в печени образуется «гемолитический фактор». В результате его идентификации была установлена липидная природа образующегося цитотоксического агента, названного липидным радиотоксином (ЛРТ). Оказалось, что ЛРТ обнаруживается уже в первые часы после облучения не только в печени, но и в крови, тонком кишечнике, семенниках, почках, желудке и других органах животных, также этот эффект был получен на растениях и микроорганизмах.ЛРТ представляет собой лабильный комплекс продуктов окисления жирных ненасыщенных кислот, гидропероксидов, эпоксидов, альдегидов и кетонов. Показано, что ЛРТ вызывает не только гемолиз, но и другие реакции, характерные для лучевого поражения: торможение клеточного деления, нарушение кроветворения, повреждение хромосомного аппарата некоторых объектов и др. В связи с этим ЛРТ был назван «естественным радиомиметиком», т. е. агентом, имитирующим биологическое действие ионизирующих излучений. Радиомиметическое действие токсических веществ липидной природы показано также в серии работ японских исследователей На рис. III.37 представлено содержание различных токсических продуктов в печени облученных крыс; видно, что в разные сроки после облучения наблюдается чаще увеличение, но иногда и уменьшение количества исследованных веществ: холина, хинонов, гистамина, продуктов автолиза, ЛРТ. Выясняя роль каждого из них, Ю. Б. Кудряшов показал, что введение ЛРТ интактным животным вызывало у них фазные изменения других «радиотоксинов», в том числе хинонов. Инъекция же хинонов или других «радиотоксинов» не приводила к образованию ЛРТ. На этом основании ЛРТ были отнесены к первичным радиотоксинам, а все остальные — к вторичным.Позднее к первичным радиотоксинам были отнесены авторами и хиноны на том основании, что и они « .способны накапливаться в клетках сразу после начала облучения как продукты первичных лучевых процессов окисления биосубстратов» Гипотеза липидных радиотоксинов базируется на выдвинутой Б. Н. Тарусовым и Н. М. Эмануэлем концепции о решающей роли в начальных процессах лучевого поражения цепных окислительных реакций свободнорадикального типа, наиболее подходящим субстратом для которых являются липиды. Так как последние представляют собой структурные элементы клеточных мембран, их поражение приводит к нарушению регуляции химизма живой клетки вплоть до уровня, приводящего ее к гибели. Возникновению цепных реакций согласно этой концепции, способствует вызываемое облучением разрушение или ингибирование природных антиокислительных (антиоксидантных) систем; в клетках, необлученных организмов они предохраняют липиды от самопроизвольного окисления.